
Footprints in the Pollen
Phil Watson
Visual and aromatic treats are only some of the pleasures provided by a carpet of wild flowers. Waiting to be discovered by the nature lover is the fascinating world of curious structures, processes and complex interrelationships between plants and animals that have evolved to ensure plants reproduce. In fact, flowers are simply the plant's tool in the mating game, with their intricate designs refined over millennia of generational change. This article explores the exciting field of pollination ecology, which studies the interaction between plants and their pollinators.
Nectar and pollen are irresistible rewards
Although flowers appear very different, they all have the same basic structure. Their central female parts (carpels) contain the ovaries which require pollinating to form fruits and seeds. These are surrounded by the male parts (stamens) which are the pollen producers, designed to ensure the fertilization of the ovules in the ovary. Encircling the stamens and pistils are the petals and sepals, colourful accessory structures whose roles are to protect the pollen and nectar and to promote their floral attributes as boldly as they dare.
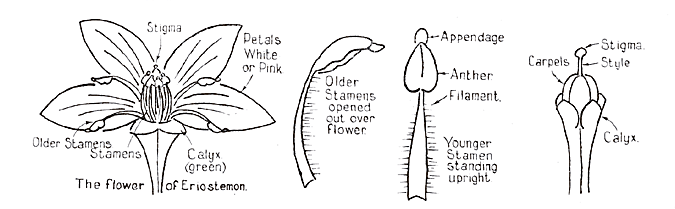 |
Parts of a Flower (Eriostemon)
From: "The Genus Eriostemon" by Jean Galbraith (Australian Plants, Vol 1 No. 4, September 1960) |
To achieve a transfer of pollen from the stamens of one flower to the ovules of another, pollination agents are targeted, attracted by a reward of delicious body building pollen which is high in fats and protein, or energy giving nectar which is high in sugary carbohydrates. To maintain a competitive advantage, the flowers' colours and scents must be as irresistible as possible. In human terms this is somewhat akin to streets lined with cake shops, cafes and restaurants displaying windows of decorated cakes and other delicacies. Like the flower-scented breezes, these are made even more tempting by fans wafting mouth watering aromas into the street.
Pollen is too expensive to waste
As there is always ample carbon in nature for plants to make carbohydrates, sugary nectar is a cheap product. In contrast, pollen demands large amounts of nitrogen and is generally at a premium in the low nitrogen environments of dry coastal heaths, grassy woodlands and vegetation communities located on siliceous and skeletal soils. In these areas nitrogen is mostly obtained through healthy symbiotic relationships between fungi and plants.
Because pollen is precious, flowers have developed structures and processes that attract pollinators while excluding other potential nectar and pollen raiders.
No footprints required
| |
 |
|
These flowers of a native violet (Viola betonicifolia) have fully opened, but often the flowers will pollinate without opening
|
Many flowers are self pollinating. Of these, a few practise the bizarre process called cleistogamony, where flowers such as Viola sp, Wahlenbergia spp, a few species of leek orchids, (Prasophyllum spp) and small-flowered Caladenia spp. pollinate without opening.
Wasteful wind and wondrous water pollination
Using wind to waft the fine pollen around is a rather wasteful and haphazard process. Typical wind pollinated plants are from the monocotyledonous families of Cyperaceae (sedges), Poaceae (grasses and cereals), Juncaceae (reeds and rushes) and a small number of dicotyledonous families including Casuarinaceae, Urticaceae and Plantagnaceae. Wind pollinated plants produce no nectar and have insignificant flowers that are mostly dioecious (separate male and female plants).
These families are responsible for many of the allergenic pollens known to induce the classic springtime hay fever and asthma attacks. But although the grasses tend to get the blame, plantains (Plantago spp.), Amaranthus sp., docks (Rumex sp.), mustards, stinging nettles and Paterson's curse are the worst culprits.
All seagrasses rely on water pollination but it is uncommon in terrestrial plants. Some wetland species such as the brilliant yellow running marsh flower Villarsia reniformis have female flowers on the water's surface which are able to attract passing pollen rafts. Radically different again is the rarely seen rain splash pollination evolved by a few plants such as large flowered Clematis.
In general most flowering plants have evolved in association with pollinators. Plants are commonly monoecious (male and female flowers on the same plant) and employ the services of diurnally active invertebrates (such as soldier beetles), marsupials and birds or nocturnally active mammals, bats and moths. Most plants recognise their own pollen and have evolved the ability for male and female parts to be receptive at different times, thus ensuring that outcrossing and hence genetic robustness is perpetuated.
Bees are nectar and pollen addicts
 |
|
Thelymitra ixioides
Spotted sun orchid
|
|
Whereas some pollinators only require nectar and/or pollen for part of their lives, bees need these honey ingredients throughout their adult and larval stages. To accumulate one kilogram of honey, bees will have pollinated a million individual flowers and flown over 50,000 kilometres.
Sadly, native bees are declining or have gradually been displaced since the introduction of the commercial honey bee and more recently the bumble bee. This has led to a reduction in native flora pollination and an increase in the pollination of weeds. Hence weeds like lupins, Agapanthus, Genista, exotic Solanum sp. and foxgloves are flourishing amongst our indigenous plants.
Recent studies of the bush pea Gompholobium huegelii report a large reduction in flower visits by the native megachilid bees where they competed with the bumble bee. The hairy body parts of this native bee perfectly match the location of the flower's anthers and stigma, whilst the equivalent areas on the bumble bee are smooth and unable to attach pollen.
In general, native bees favour the blue and purple flowered sun orchids (Thelymitra spp.), blue iris (Patersonia sp.), blue bush pea (Hovea sp.) and 'Happy Wanderers' (Hardenbergia sp.) although they are happy to add their footprints in the pollen of yellow, mauve, pink and white blooms.
Pollination by long tongues, short tongues or buzzing wings
Bees are capable of manipulating a variety of floral constructions to gain access to the nectaries. Some flowers have developed finely tuned relationships so that only a few species can decode the cryptic clues leading them to the flowers' rewards. A classic example is the bloodroot, Haemodorum sp., where specialised bees are able to enter and pollinate their closed flowers, thus eliminating other nectar competitors.
| |
 |
|
 |
|
Left: Haemodorum coccineum, Scarlet bloodroot.
Right: Epacris longiflora, Fuchsia heath
|
The largest and most commonly seen bees are the sparsely hairy short-tongued species that have evolved in parallel with the Myrtaceae family. Gums (Eucalyptus spp.), tea trees (Leptospermum spp.), paperbarks (Melaleuca spp.) and heath myrtles (Baeckea spp.) have shallow cup-like nectaries ideally suited for lapping by short tongues. Prolific nectar producers such as native box, Bursaria spinosa, and waxflowers (Philotheca spp.) are also suited for short tongued bees.
Only a small number of the estimated 700 species of solitary bees are long-tongued. One, the leaf-cutter bee, confines foraging to flowers with deep floral tubes such as native fuchsia (Correa spp.), native rosemary (Westringia sp.) and native heath (Epacris spp.). As they feed, pollen is dusted onto their bodies or hairy legs and carried to the stigma of another flower. This process has recently been short circuited by the ubiquitous bumble bee having learnt to steal the nectar by drilling into the base of the epacrid's corolla tube.
Of horticultural importance to commercial growers for pollinating glass house tomatoes is the bumble bee's 'buzz pollination technique'. However, it is also feasible that native buzz pollinators such as the large, hairy, metallic green Carpenter Bee and the smaller Blue Banded Bee could fulfil this role. These bees collect the pollen that is flung from vibrating flower anthers. They carry it to nesting holes bored in the soft wood of Grass-trees' (Xanthorrhoea) flower shafts or old banksia trunks. Here, an egg in each cell is provided with a feed of pollen and nectar porridge before closing off with wax and frass.
Species such as the golden Guinea flower, Hibbertia riparia, kangaroo apple, Solanum laciniatum, Fringe lilies (Thysanotus spp.) and flax lilies, Dianella spp, rely on buzz pollination but are often inundated by ineffective raiders such as honey bees. Interestingly, Hibbertia spp. have no nectary, but instead rely on their pollen to attract those bees that are searching for high protein food to feed their larva.
Members of the Goodeniaceae family such as native primrose, Goodenia lanata, and spur velleia, Velleia paradoxa, also have buzz-pollinated flowers, but they are designed differently to protect them from self pollinating. With the aid of a cup-like structure (indusium) they catch maturing pollen that is ready for dispersal. Once the pollen is dispersed or becomes non viable, the stigma will change to become receptive for pollination. Most Goodeniaceae have the nectary spur hidden at the base of the petals which restricts nectar access to long tongued pollinators.
Bush peas are a bee's favourite
The bush peas (Fabaceae) with their functionally designed wing and keel structure (e.g. showy bossiaea, Bossiaea cinerea, bitter peas, Daviesia sp., and golden pea, Aotus ericoides) along with the pea-like flowers of the Polygalaceae (e.g. blue love creeper, Comesperma volubile) have evolved an ingenuous system suited for bee pollination. With the anther and stigma protected below the flexible lower keel petal, only appropriately sized bees can part the petals and trip the spring loaded organs to tickle the bee's hairy belly.
Nectar Guides
 |
|
Pelagonium australe
Austral stork's bill
|
|
Intriguing associations have evolved with native bees and other insects whose eye receptors are strongest in the far blue end of the UV spectrum. This allows flowers that appear white to mammals and birds to display distinctive UV colour patterns which act as nectar guides. Common UV patterns include light blues with darker maroon or violet streaks and patches.
Coupled with the flower's ability to emit romantic pheromone attractants, the guides become magnets for bees and other insects. Numerous examples include the vivid set of spots and streaks on donkey orchids (Diuris spp.) and bush peas, the rosy red streaks on the petals of native pelagonium, Pelagonium australe and the prominent purple veins on the floral throats of native flax, Linum marginale.
The bee and orchid love story
| |
 |
|
Diuris sulphurea
Tiger orchid
|
Due to the lack of a nectary, the pollination of many native terrestrial orchids is contingent on their ability to mimic nectar-bearing flowers found within their vegetation communities. (The tiger orchid, Diuris sulphurea, and leopard orchid, D.paradina, mimic numerous bush peas by replicating their structure, colours and timing of opening.) This, coupled with their ability to emit the female bee's pheromones, lures the male bee into attempting to mate with the labellum, which results in pollen transfer.
Similarly, blue-flowered species such as sun orchids (Thelymitra spp.), small flowered caladenias and wax lip orchid (Glossodia sp.), have modified labella that are similar in size and shape to their petals.
This has enabled them, with their strong aromatic scents, to mimic blue flowered irises and/or lilies (eg. blue stars, Cbamaescilla) and attract a similar suite of pollinating insects, mostly native bees.
Pollen Clogging
As insects learn to recognise the flowers that provide key food, the colour, shape and overall appearance of the flower are soon retained in their memory. Specialised native bees, like many other insects, indiscriminately visit flowers that fit within the same group or 'guild' (e.g. blue lilies and sun orchids). Hence they often accumulate pollen on their bodies from several different species. With this mix of pollen types scattered over their bodies, foreign pollen can be inadvertently placed on the stigma of a flower preventing pollen of its own species reaching the stigma. This 'pollen clogging' is cunningly avoided - the anthers position the pollen consistently on one point of the bee's hairy body, so that only the perfectly poised stigma receives a brushing of this pollen.
The many other Pollinators
During peak flowering season, wildflowers are enveloped in clouds of pollinating insects such as flies, wasps, gnats, beetles, moths and butterflies, all busily leaving their footprints in the pollen. There are other plant-insect interrelationships, and birds and mammals also contribute to pollination ecology. But that is another story!
Recommended Reading
Buchmann,S.L. & Nabhan, G.P.,1996, The Forgotten Pollinators; Island Press, USA.
Faegri, K. & Van der Pijl, 1973, The Principles of Pollination Ecology, Collins, London.
Harvey, M.S. & Yen A.L., 1997, Worms to Wasps: An illustrated Guide to Australia's Terrestrial Invertebrates, Oxford University Press, Melbourne.
Kirkpatrick, J.B. & Harris, S., 1999, The Disappearing Heath Revisited, Tasmania Environment Centre Inc., Hobart.
Nevill, Simon, 2001, Guide to Wildflowers of South Western Australia, Simon Nevill Publications.
From "Eucryphia", the newsletter of the Australian Plants Society (Tasmania), July 2007.
Australian Plants online - 2007
Association of Societies for Growing Australian Plants
|